ODDYCHANIE BEZTLENOWE

Substratem oddychania beztlenowego najczęściej jest glukoza. Tutaj przemiany pirogronianu polegają na jego bezpośredniej (fermentacja mlekowa) lub pośredniej redukcji (fermentacja alkoholowa), której towarzyszy utlenienie NADH+H+ do NAD. W łańcuchu oddechowym pirogronian nie może być dalej utleniany i dlatego tak się dzieje. Fermentacji nadaje się nazwę w zależności od produktu końcowego.
Fermentacja mlekowa zachodzi w komórkach bakterii mlekowych, u pasożytów wewnętrznych i mięśniach szkieletowych, gdy brakuje tlenu. Końcowym produktem tej fermentacji jest mleczan. Powstający w mięśniach mleczan jest transportowany z krwią do wątroby, a następnie ulega przemianie w glukozę w procesie glukoneogenezy. Glukoza wraca z krwią do mięśni, gdzie służy jako substrat do odbudowy zapasów glikogenu.
Fermentacja alkoholowa przebiega w komórkach drożdży, w nasionach okrytych twardą łupiną, korzeniach, dużych mięsistych owocach. Powstały w glikolizie pirogronian ulega dekarboksylacji do aldehydu octowego, a następnie do etanolu.
ODDYCHANIE TLENOWE
Jest znacznie bardziej skomplikowane. Spróbuję wypisać je w punktach:
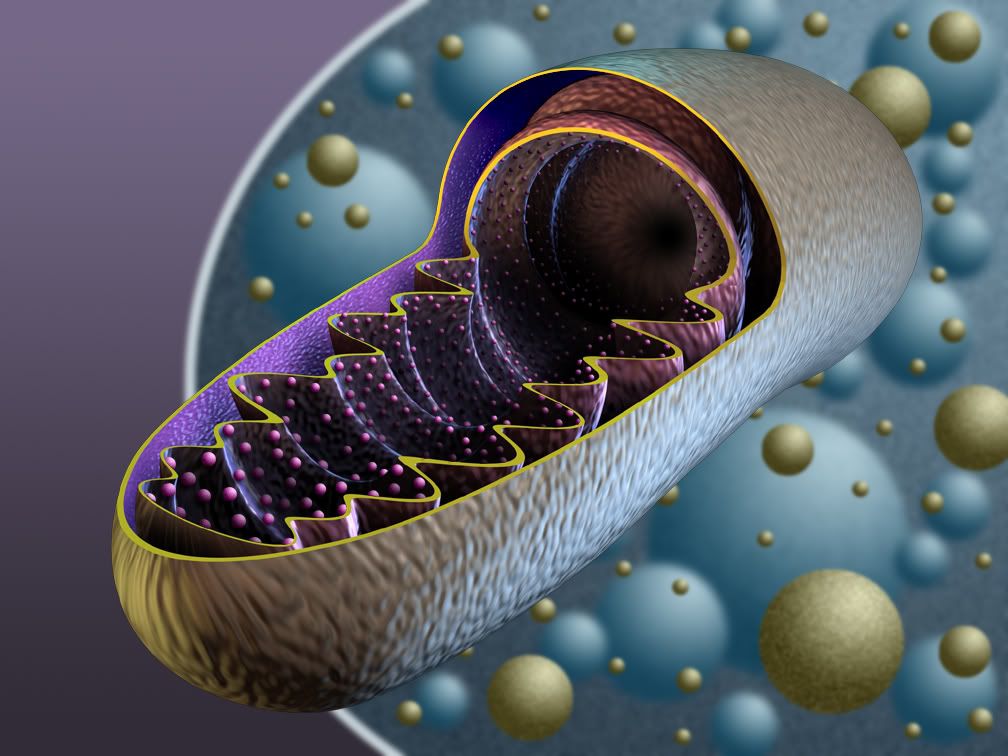
1. Powstający w glikolizie pirogronian wędruje do mitochondriów, gdzie ulega oksydacyjnej dekarboksylacji do związku dwuwęglowego - aktywnego octanu, zwanego acetylokoenzymem A, wiec inaczej acetylo- Co A.
2. Acetylo-Co A zostaje włączony w cykl przemian kwasów karboksylowych zwany cyklem Krebsa lub cyklem kwasu cytrynowego. Tam zostaje utleniony do dwóch cząsteczek CO2.
3. Akceptorem aktywnego octanu w cyklu Krebsa jest 4-węglowy szczawiooctan. W wyniku reakcji kondensacji powstaje 6-węglowy cytrynian, od którego pochodzi nazwa cyklu.
4. Przez szereg produktów pośrednich zostaje odtworzony związek 4-węglowy, który przyłącza kolejną cząsteczkę acetylokoenzymu A.
5. Na czterech etapach cyklu odbywa się proces utleniania poprzez odwodorowanie z wydzieleniem trzech cząstek NADH+H+ i FADH2. Podczas jednego pełnego cyklu Krebsa dochodzi do dwukrotnej dekarboksylacji i 4-krotnego odwodorowania.
6. Zredukowane przenośniki przenoszą wodór na błonę grzebieni mitochondrialnych, gdzie zlokalizowany jest łańcuch oddechowy.
7. Łańcuch oddechowy to ciąg oksyreduktaz: NAD, FAD, ubichinon, cytochrom b, cytochrom c, oksydaza cytochromowa.
8. Na początku łańcucha płynie strumień atomów wodoru, a następnie przez wyższy potencjał oksydoredukcyjny strumień elektronów.
9. W łańcuchu oddechowym produkowane jest ATP. Ostatecznym akceptorem elektronów i protonów wodorowych jest tlen w związku z czym produkcję ATP w łańcuchu oddechowym określamy jako fosforylację oksydacyjną.

Substratem oddychania beztlenowego najczęściej jest glukoza. Tutaj przemiany pirogronianu polegają na jego bezpośredniej (fermentacja mlekowa) lub pośredniej redukcji (fermentacja alkoholowa), której towarzyszy utlenienie NADH+H+ do NAD. W łańcuchu oddechowym pirogronian nie może być dalej utleniany i dlatego tak się dzieje. Fermentacji nadaje się nazwę w zależności od produktu końcowego.
Fermentacja mlekowa zachodzi w komórkach bakterii mlekowych, u pasożytów wewnętrznych i mięśniach szkieletowych, gdy brakuje tlenu. Końcowym produktem tej fermentacji jest mleczan. Powstający w mięśniach mleczan jest transportowany z krwią do wątroby, a następnie ulega przemianie w glukozę w procesie glukoneogenezy. Glukoza wraca z krwią do mięśni, gdzie służy jako substrat do odbudowy zapasów glikogenu.
Fermentacja alkoholowa przebiega w komórkach drożdży, w nasionach okrytych twardą łupiną, korzeniach, dużych mięsistych owocach. Powstały w glikolizie pirogronian ulega dekarboksylacji do aldehydu octowego, a następnie do etanolu.
ODDYCHANIE TLENOWE
Jest znacznie bardziej skomplikowane. Spróbuję wypisać je w punktach:
1. Powstający w glikolizie pirogronian wędruje do mitochondriów, gdzie ulega oksydacyjnej dekarboksylacji do związku dwuwęglowego - aktywnego octanu, zwanego acetylokoenzymem A, wiec inaczej acetylo- Co A.
2. Acetylo-Co A zostaje włączony w cykl przemian kwasów karboksylowych zwany cyklem Krebsa lub cyklem kwasu cytrynowego. Tam zostaje utleniony do dwóch cząsteczek CO2.
3. Akceptorem aktywnego octanu w cyklu Krebsa jest 4-węglowy szczawiooctan. W wyniku reakcji kondensacji powstaje 6-węglowy cytrynian, od którego pochodzi nazwa cyklu.
4. Przez szereg produktów pośrednich zostaje odtworzony związek 4-węglowy, który przyłącza kolejną cząsteczkę acetylokoenzymu A.
5. Na czterech etapach cyklu odbywa się proces utleniania poprzez odwodorowanie z wydzieleniem trzech cząstek NADH+H+ i FADH2. Podczas jednego pełnego cyklu Krebsa dochodzi do dwukrotnej dekarboksylacji i 4-krotnego odwodorowania.
6. Zredukowane przenośniki przenoszą wodór na błonę grzebieni mitochondrialnych, gdzie zlokalizowany jest łańcuch oddechowy.
7. Łańcuch oddechowy to ciąg oksyreduktaz: NAD, FAD, ubichinon, cytochrom b, cytochrom c, oksydaza cytochromowa.
8. Na początku łańcucha płynie strumień atomów wodoru, a następnie przez wyższy potencjał oksydoredukcyjny strumień elektronów.
9. W łańcuchu oddechowym produkowane jest ATP. Ostatecznym akceptorem elektronów i protonów wodorowych jest tlen w związku z czym produkcję ATP w łańcuchu oddechowym określamy jako fosforylację oksydacyjną.












.jpg)






















_3D_sm.png) -
- 

 .
.

