 Organizmy autotroficzne, charakteryzujące się prostą budową lub żyjące w środowisku wodnym pobierają wodę całą powierzchnią ciała. Dla roślin lądowych źródłem wody są opady atmosferyczne. Rośliny wodne są w stanie pełnego uwodnienia, dlatego nie wykazują wyraźnej siły ssącej. Większość roślin lądowych pobiera wodę z gleby przez korzenie. W strefie włośnikowej jest znacznie powiększona powierzchnia chłonna.
Organizmy autotroficzne, charakteryzujące się prostą budową lub żyjące w środowisku wodnym pobierają wodę całą powierzchnią ciała. Dla roślin lądowych źródłem wody są opady atmosferyczne. Rośliny wodne są w stanie pełnego uwodnienia, dlatego nie wykazują wyraźnej siły ssącej. Większość roślin lądowych pobiera wodę z gleby przez korzenie. W strefie włośnikowej jest znacznie powiększona powierzchnia chłonna.
MECHANIZM BIERNY opiera się na sile ssącej liści powstającej na skutek transpiracji, czyli parowania wody z liści. Wyparowywanie wody z powierzchniowej warstwy komórek liści powoduje zmniejszenie ich potencjału wody i turgoru, a wzrost siły ssącej. Wyparowywanie wody sięga przez łodygę do korzenia na skutek czego woda przechodzi z korzeni do naczyń i cewek, a z gleby do włośników. W ten sposób jest zapewniona możliwość transportu wody do wierzchołka korony najwyższych drzew. W tym wypadku liście są sprawcami pobierania i transportu wody, natomiast korzenie pełnią funkcję jedynie bierną. Mechanizm ten nie wymaga dostarczenia energii (poza energią słoneczną) i to jest plus, jednak wadą jego jest to, że jest skuteczny jedynie w warunkach sprawnej transpiracji (w innym wypadku potrzebny jest transport aktywny np. wiosną, bo pączki są słabo rozwinięte lub nocą)

SIŁA KOHEZJI również ma wpływ na ruch wody w naczyniach. Jeśli zanurzymy w szklance rurkę szklaną, to dzięki sile kohezji woda podniesie się w niej ponad poziom wody w pojemniku. Im cieńsza rurka, tym wyżej się podniesie, a przecież naczynia roślin są bardzo cienkimi rurkami.

Siła kohezji na przykładzie przyciągających się tancerzy,
istnieje siła przyciągających się do siebie na środku, ale również
tych do ściany, dlatego im cieńsza rurka tym bardziej idą w górę
MECHANIZM AKTYWNY

Opiera się na aktywnym transporcie jonów przez błonę komórkową roślin. Proces ten jest związany ze zużyciem ATP. Woda przenikając z gleby do komórek ryzoderm i dalej, w poprzek korzenia do naczyń i cewek powoduje wzrost ciśnienia w elementach przewodzących (poziom wody się podnosi). To wywierane ciśnienie nazywa się parciem korzeniowym. Jego siła jest niższa od siły ssącej liści.
TRANSPIRACJA

Największą powierzchnię parowania tworzą liście. Wydzielanie wody w postaci pary przez rośliny lądowe nosi nazwę transpiracji.
Szybkość parowania jest tym większa, im wyższa jest temperatura, im większa jest powierzchnia swobodna cieczy i im mniej jest pary w powietrzu ponad cieczą.
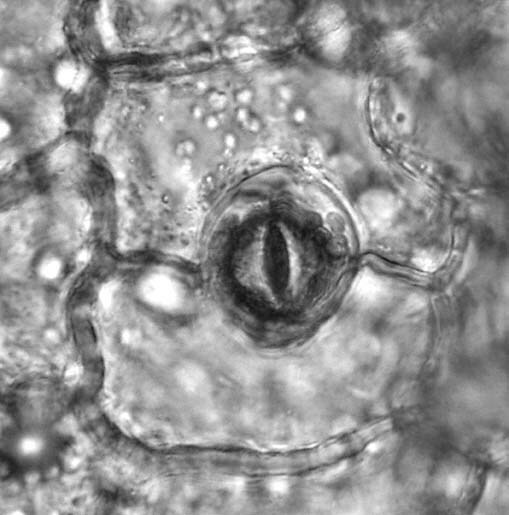
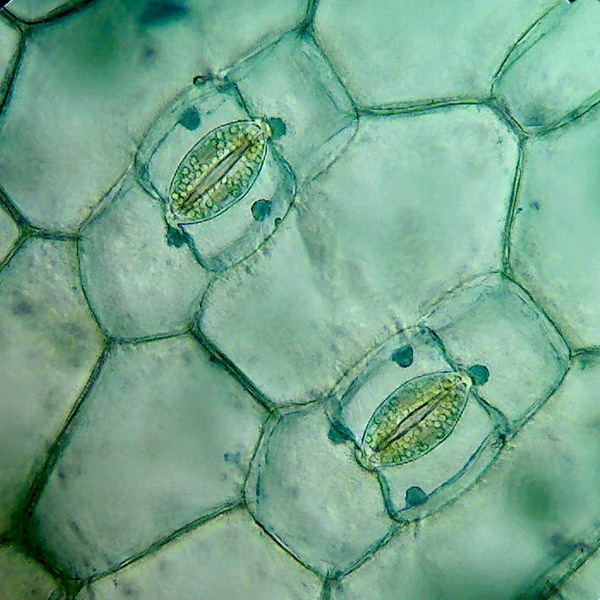
Wyróżnia się transpirację kutikularną, która zachodzi bezpośrednio przez skórkę oraz transpirację szparkową, która odbywa się przez aparaty szparkowe (ta jest bardziej efektywna)
Stopień rozwarcia aparatów szparkowych nie zawsze wpływa na intensywność transpiracji. Główną przyczyną ograniczenia transpiracji może być w nocy brak dopływu do rośliny energii świetlnej zamienianej na ciepło, ponieważ transpiracja zużywa ok. 80% energii świetlnej pochłoniętej przez liść.
U roślin iglastych i jednoliściennych, które mają niewielką powierzchnię liścia, aparaty szparkowe rozmieszczone są na obu stronach liścia. U roślin dwuliściennych są umieszczone na spodniej stronie, co ogranicza nadmierne parowanie i utratę wody. U roślin wodnych aparaty szparkowe umieszczone są po wierzchniej stronie blaszki liściowej.
 Aby transpiracja mogła zachodzić aparaty szparkowe muszą być otwarte. Ruch tych aparatów zachodzi przy działaniu różnych mechanizmów. Aktywny transport jonów potasowych do komórek szparkowych powoduje wzrost uwodnienia, a w efekcie wzrost ciśnienia turgorowego, to z kolei prowadzi do otwarcia aparatu szparkowego. W nocy zaś przemiana glukozy w skrobię również wpływa na ciśnienie turgorowe. W dzień na odwrót skrobia jest zamieniana w glukozę.
Aby transpiracja mogła zachodzić aparaty szparkowe muszą być otwarte. Ruch tych aparatów zachodzi przy działaniu różnych mechanizmów. Aktywny transport jonów potasowych do komórek szparkowych powoduje wzrost uwodnienia, a w efekcie wzrost ciśnienia turgorowego, to z kolei prowadzi do otwarcia aparatu szparkowego. W nocy zaś przemiana glukozy w skrobię również wpływa na ciśnienie turgorowe. W dzień na odwrót skrobia jest zamieniana w glukozę.
Dzięki transpiracji powstaje siła ssąca liści umożliwiająca przepływ wody przez roślinę bez wydatkowania energii z ATP. Przyczyniając się jednak do utraty wody, ogranicza wzrost i rozwój rośliny.
TRANSPORT BLISKI - pomiędzy sąsiednimi komórkami, TRANSPORT DALEKI to przepływ wody z solami mineralnymi i asymilatów pomiędzy organami roślin. Rodzaje transportu dalekiego:
- transport wody z solami mineralnymi odbywa się w cewkach i naczyniach z dołu do góry
- transport związków organicznych odbywa się w komórkach sitowych, w różnych kierunkach, w zależności od potrzeb.
TRANSPORT ASYMILATÓW
Postacią transportową asymilatów (produktów fotosyntezy) u roślin jest sacharoza. Powstaje ona w komórkach donora (ten który dostarcza, dawca), czyli miękiszu asymilacyjnego i jest aktywnie pompowana (z użyciem ATP) do rurek sitowych, co nazywa się załadunkiem floemu. Sacharoza obniża potencjał wodny rurek sitowych powodując napływ wody, następnie komórki akceptora przepompowują sacharozę z floemu do własnych komórek, gdzie jest ona zamieniana w skrobię nieaktywną osmotycznie. To nazywamy rozładunkiem floemu.
INNE WAŻNE INFORMACJE:
- włośniki pobierają wodę z gleby dzięki niższemu potencjałowi wody niż w roztworze glebowym. Do naczyń lub cewek płynie ona kanałem symplastycznym, czyli z komórki do komórki w cytozolu lub kanałem apoplastycznym - w przestworach międzykomórkowych.
- roślina wybiórczo pobiera jony (czyli tylko te, których potrzebuje)
PYTANIE MATURALNE:
Źródło: WSiP

Opiera się na aktywnym transporcie jonów przez błonę komórkową roślin. Proces ten jest związany ze zużyciem ATP. Woda przenikając z gleby do komórek ryzoderm i dalej, w poprzek korzenia do naczyń i cewek powoduje wzrost ciśnienia w elementach przewodzących (poziom wody się podnosi). To wywierane ciśnienie nazywa się parciem korzeniowym. Jego siła jest niższa od siły ssącej liści.
TRANSPIRACJA
Największą powierzchnię parowania tworzą liście. Wydzielanie wody w postaci pary przez rośliny lądowe nosi nazwę transpiracji.
Szybkość parowania jest tym większa, im wyższa jest temperatura, im większa jest powierzchnia swobodna cieczy i im mniej jest pary w powietrzu ponad cieczą.
Wyróżnia się transpirację kutikularną, która zachodzi bezpośrednio przez skórkę oraz transpirację szparkową, która odbywa się przez aparaty szparkowe (ta jest bardziej efektywna)
Stopień rozwarcia aparatów szparkowych nie zawsze wpływa na intensywność transpiracji. Główną przyczyną ograniczenia transpiracji może być w nocy brak dopływu do rośliny energii świetlnej zamienianej na ciepło, ponieważ transpiracja zużywa ok. 80% energii świetlnej pochłoniętej przez liść.
U roślin iglastych i jednoliściennych, które mają niewielką powierzchnię liścia, aparaty szparkowe rozmieszczone są na obu stronach liścia. U roślin dwuliściennych są umieszczone na spodniej stronie, co ogranicza nadmierne parowanie i utratę wody. U roślin wodnych aparaty szparkowe umieszczone są po wierzchniej stronie blaszki liściowej.
Dzięki transpiracji powstaje siła ssąca liści umożliwiająca przepływ wody przez roślinę bez wydatkowania energii z ATP. Przyczyniając się jednak do utraty wody, ogranicza wzrost i rozwój rośliny.
TRANSPORT BLISKI - pomiędzy sąsiednimi komórkami, TRANSPORT DALEKI to przepływ wody z solami mineralnymi i asymilatów pomiędzy organami roślin. Rodzaje transportu dalekiego:
- transport wody z solami mineralnymi odbywa się w cewkach i naczyniach z dołu do góry
- transport związków organicznych odbywa się w komórkach sitowych, w różnych kierunkach, w zależności od potrzeb.
TRANSPORT ASYMILATÓW
Postacią transportową asymilatów (produktów fotosyntezy) u roślin jest sacharoza. Powstaje ona w komórkach donora (ten który dostarcza, dawca), czyli miękiszu asymilacyjnego i jest aktywnie pompowana (z użyciem ATP) do rurek sitowych, co nazywa się załadunkiem floemu. Sacharoza obniża potencjał wodny rurek sitowych powodując napływ wody, następnie komórki akceptora przepompowują sacharozę z floemu do własnych komórek, gdzie jest ona zamieniana w skrobię nieaktywną osmotycznie. To nazywamy rozładunkiem floemu.
INNE WAŻNE INFORMACJE:
- włośniki pobierają wodę z gleby dzięki niższemu potencjałowi wody niż w roztworze glebowym. Do naczyń lub cewek płynie ona kanałem symplastycznym, czyli z komórki do komórki w cytozolu lub kanałem apoplastycznym - w przestworach międzykomórkowych.
- roślina wybiórczo pobiera jony (czyli tylko te, których potrzebuje)
PYTANIE MATURALNE:
Źródło: WSiP
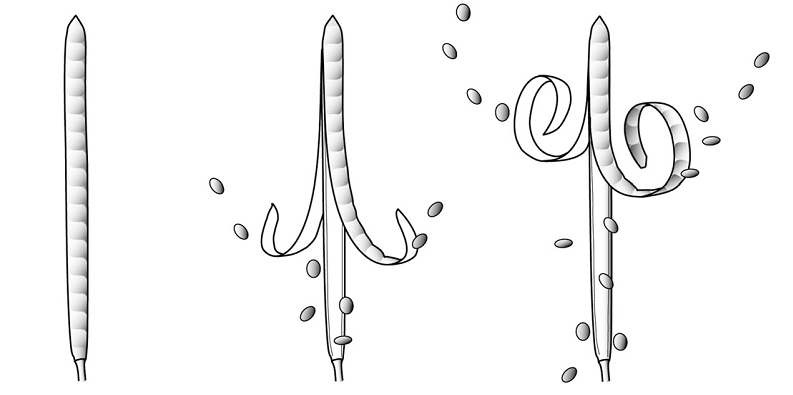
Na schemacie przedstawiono kierunki transportu różnych substancji w organizmie rośliny nasiennej.
a. Wyjaśnij, dlaczego transport asymilatów zachodzi zarówno w górę, jak i w dół rośliny.b. Podaj nazwę procesu, w którego wyniku woda opuszcza organizm rośliny.
























.jpg/250px-Resin_with_insect_(aka).jpg)









 Kolce róży
Kolce róży


